,
The algal bloom and climate anomalies
of 1992 Dr W J Ballantine Leigh Laboratory Seminar, May 28th, 1993
[this paper has been reproduced here for those
wishing to study
the plankton blooms of 1992 and their effects
on the environment.
To our knowledge it is not available elsewhere.
- Floor Anthoni.
Please note that this is not an official scientific
publication!]
Use your BACK button to reverse out of this
document. www.seafriends.org.nz/enviro/plankton/bloom92.htm
.
'
Introduction A the end of August last year an intense microalgal bloom developed
along the open coast at the Leigh Laboratory. This bloom was unusual in
its intensity, seasonal timing, duration, geographic extent, composition
and subsequent biological events. This seminar attempts to review the data
about this bloom and discuss implications. It is unusual in several respects:
It reports on the work of many people including:
Allison Haywood, MSc Environmental Sciences: studies of particles
in open water
Leslie Rhodes, Cawthron Institute: identification and counts of
microalgae
ARC and others: water quality sampling
Jo Evans and others: local climate data
Meteorological services: satellite derived data and climate indices
Many biologists at Leigh: anomalous biological data since Sept 1992
Fishermen and divers: reports of seawater discoloration and low
underwater visibility.
There was no formal project. No one had the task of investigating the bloom
as such, or of assigning anyone to do so. Those who involved themselves
all had different aims and interests, and none included a direct study
of the bloom.
It raises not only the standard scientific questions on the bloom itself
- what was its nature, causes and consequences - but also wider
questions such as:
what is the proper scientific attitude to rare but significant events
in your general area of interest, but outside your current range of concern?
what are the proper methods for combining a wide range of data which
vary widely in reliability, and scales of time, space and complexity?
are there any general principles which would help, or are we obliged
to react each time without any guidelines?
These questions are made more pointed because of:
the current policies for 'user pays' and competitive research
the contrast between professional reactions to the algal blooms of 1992
and the toxic shellfish issues of 1993.
1. The intensity, duration and extent of the bloom Evidence at Leigh
Algal pigment, total seston and turbidity sampling by Alison Haywood.
Counts, cultures and identifications of algae by Leslie Rhodes, Cawthron.
Diver observations of underwater visibility
Routine observations of seawater colour from clifftop (during daily tea
breaks!)
Evidence from further afield
Aerial survey by Alison Haywood of the western Hauraki Gulf, September
13th
Counts and identifications from inner W. Gulf and Coromandel by L. Rhodes.
Personal reports from fishermen and divers on water colour and underwater
visibility
Newspaper reports on water colour, etc.
Results and conclusions
There is no routine monitoring of phytoplankton anywhere in the region,
and the marine water quality parameters that are monitored are unsuitable
to estimate even gross phytoplankton abundance.
Previous studies of phytoplankton in the region have produced data of high
scientific value, but they used such varied methods and standards that
the data are rarely comparable.
Chlorophyll-a concentration near the Leigh Laboratory increased tenfold
at the end of August and remained high until this sampling finished at
the end of October (from a mean 0.8 µg/l to mean 9.2 µg/l).
This compares with 1986 mean values of 0.9 µg/l for winter and 3.1
µg/l for spring, in roughly the same area (Booth and Sonergaard,
1989)
Over the same period, much smaller increases (approximately doubling) were
recorded in the dry weight of total seston and in measurements from a standard
turbidity meter (which measures scatterance rather than absorbance of light).
From 8th October (first sample) through November, counts of phytoplankton
in water samples from the same site at Leigh showed high to very high abundances
of several flagellate microalgae. By the end of December samples were more
or less normal for the time of year.
The periods of high algal pigments and/or high algal counts coincided with
unusually coloured seawater which was sufficiently obvious to be noticed
by divers, observers in boats and from the cliff-top.
On September 13th, an aerial survey from Auckland to Mangawhai Heads showed
strongly discoloured water extending over virtually the whole western half
of the Hauraki Gulf.
On several occasions during September-November, at the entrance to the
Whangateau Harbour, the incoming tide consisted of discoloured water of
very low underwater visibility (1-2m), while the outgoing tide was much
clearer (5-7m visibility).
During September through November many reports from fishermen indicated
that strongly discoloured seawater occurred for long periods over virtually
the whole Hauraki Gulf west of a line joining Bream Head to Coromandel,
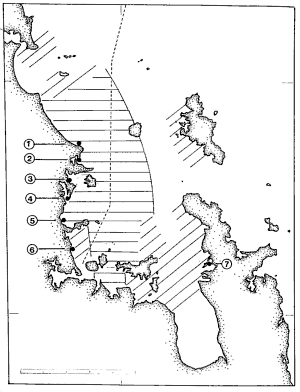
with more variable occurrences further north and east.
All reports were consistent with:
strongly discoloured water (varying in intensity by the day) over most
of the inner and western Gulf north to Leigh
usually discoloured water but banded in extent and/or intermittent in time
extending from the Hen and Chickens, through Little Barrier to Coromandel,
and off western Gt Barrier.
patches or bands of discoloured water at times at the Poor Knights, off
Whangarei Heads and at Mokohinau.
The evidence from further afield is very poor. Scattered reports from the
Bay of Islands, Bay of Plenty and South Island suggest that unusual coloured
water did occur but it is not clear whether this was atypical in timing
or intensity.
2. The timing and species composition of the bloom
Evidence:
Whole water samples counted by Leslie Rhodes (Cawthron Inst):
Ti Point to Browns Bay (5 sites) and Coromandel (3 sites) in November only.
Filter samples analysed by Allison Haywood: 2-3 times weekly from late
May to October and daily during September.
Various published and thesis accounts of phytoplankton in the Hauraki Gulf
in earlier years.
Results and conclusions:
The bloom commenced
off the Leigh Laboratory at the end of August. It remained intense through
November and declined to more normal levels and composition in December.
The bloom was dominated by three species:
A rhaphidophyte Fibrocapsa japonica (appr 30µm dia)
A silicoflagellate Dictyocha speculum (appr 40µm dia)
A prymnesiophyte Gephyrocapsa oceanica (appr 10µm dia)
N.B. By their usual size definitions, Fibrocapsa and Dictyocha
are 'net' phytoplankton (20-200µm) but neither are likely to be recorded
by net sampling and may not show in preserved samples. Gephyrocapsa
is classed as 'nanoplankton' (2-20µm)
There are problems with identification. First the species concept is
of doubtful meaning at this level. There are few morphological characteristics,
many of these are hard to detect in light microscopy, or are known to vary
within the 'species'. Critical 'identification' involves culturing and
comparing with known 'strains' and/or the use of electron microscopy (EM).
Second, the original samples are very small compared to the sea and further
sub-sampling is necessary for counts. Only a few specimens are selected
for EM examination or culture. Third, very little work has been done on
many of these 'species'.
Although diatoms and dinoflagellates were present throughout the bloom,
and these were generally of the expected species, they formed a small to
very small proportion of the total phytoplankton observed.
The contribution of picoplankton (0.2-2µm) to the biomass of the
bloom is unknown (no checks or counts known for any site during the bloom),
but may have been significant. Booth and Sonergaard (1989) sampling the
Hauraki Gulf (mainly near Leigh) in 1986 found picoplankton proportionally
more in winter (mean 33% of total chlorophyll-a) but absolutely more abundant
in spring.
In early October the bloom was dominated (in terms of cell volumes) by
Fibrocapsa
japonica, but later the armoured form of Dictyocha speculum
was co-dominant. The small coccolithophorid Gephyrocapsa oceanica
was abundant throughout the bloom, but was sub-dominant in biomass.
Previously published data for the Leigh region is hard to relate to the
1992 results because of the variety of sampling systems, taxonomic selection,
and analysis. Nevertheless standard sampling and analysis (i.e. nets, hoses
or dipped water, then identifications and counts) never reported these
or similar species as biomass dominants. Furthermore the biomass reached
in 1992 is unique apart from the summer bloom of 1983.
Fibrocapsa was not previously reported at all and
Dictyocha
(while noted as present) was never regarded as being worth counting. A
coccolithophorid, identified as Emiliana huxleyi (but which almost
certainly was Gephyrocapsa oceanica) was reported as present each
spring for a short burst.
The 'spring blooms' previously reported seem to be variable
in amount, timing and composition (between years), but mostly consist of
an increase in diatom numbers together with an increase in their proportion
of total phytoplankton biomass.
3. The physical background
Evidence
Leigh Laboratory daily records 1967 to present, including sea surface temperature
(SST), rainfall, solar radiation, wave surge, wind direction and run.
Satellite-derived sea surface temperature anomalies.
Southern Oscillation Index (SOI) calculations.
Conclusions:
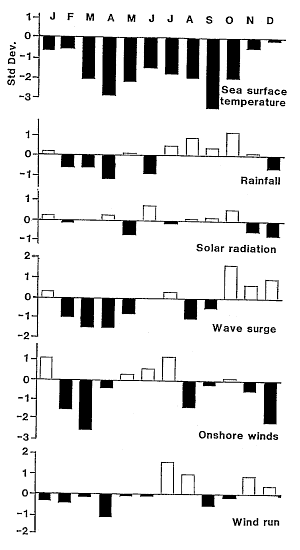
Sea surface temperatures
(SST) were at their lowest when the bloom commenced but solar radiation
had increased markedly some two weeks before.
During 1992 all climate factors (except SST) were not especially different
from average or their variations were unrelated to the development of the
algal bloom. Sea temperatures, however, were markedly colder than usual
for six months prior to the bloom. The onset of the bloom immediately followed
the coldest sea temperatures ever recorded at Leigh.
Reliable daily salinities are not available for 1992, due to a failure
in instrument calibration, but the available data suggests that salinity
was close to or slightly below normal during July through October. Records
from the Japanese research vessel Hakuho-maru which passed through
the area on October 7th 1992, show normal salinities for the season, i.e.
>35.0 PSU north of Leigh and <34.9 PSU to the south. There was no evidence
of any stratification in the Leigh area at least until November.
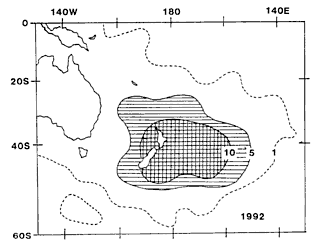
Satellite-derived data of SST anomalies, produced by the Australian Bureau
of Meteorology as monthly maps, shows that the highly anomalous sea temperatures
recorded at Leigh were part of a widespread pattern (centred east of New
Zealand and encompassing the whole country) which lasted throughout most
of 1992.
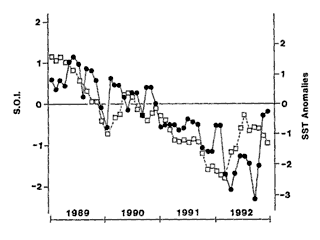
The SST anomalies of 1992 continued a downward trend from the previous
3 years. 1989 was very arm, 1990 near average, and 1991 was very cool.
This 4 year trend closely followed a trend in the Southern Oscillation
Index (SOI), which signals the El Niño/La Niña changes of
the topical Pacific.
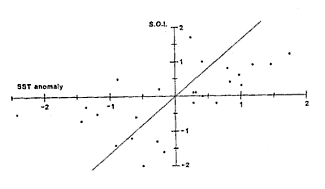
There is a clear trend relating SST anomalies at Leigh to the SOI (p=0.014)
but it only explains 35% of the variance.
4. The triggering, limiting and forcing factors
While the precise
triggers for the increase of particular species involved in the bloom and
the limiting factors affecting their subsequent abundance are likely to
vary both specifically and regionally, the widespread nature of the bloom
and its duration suggest some general forcing factor.
The most obvious candidate is the extremely low, widespread and long-lasting
sea surface temperature anomalies. These could have acted in several ways
- e.g. to inhibit the normal phytoplankton community, to encourage cold-adapted
species, to inhibit grazers or any combination of these.
The idea of a widespread climatic forcing factor is supported by the total
set of biological patterns, especially those related to frequency of occurrence.
The SST anomalies of 1992 were rare and extreme events, indeed unique in
the 26 year record at Leigh. The algal blooms were also rare and extreme,
both in themselves and in the subsequent biological disruptions.
The unusual features of the bloom included not just its intensity, but
also its duration, its geographic extent, the species involved, their multiple
dominance and their regional differences.
The coincident and subsequent biological disruptions (see next section)
included a wide range of habitats and groups, as well as similar geographic
extent.
The nearest equivalent climatic event at Leigh was the summer of 1983.
The very low sea temperatures then were also accompanied by a major, widespread
algal bloom and severe biological disruptions, although the season was
opposite and the species involved were generally different from those of
1992 (Taylor et al, 1985)
In both cases, while the detailed causes and limiting factors are likely
to have been highly variable, the very low sea temperatures appear to have
acted as a general forcing factor.
It is widely assumed that since (a) nutrients must quickly limit the exponential
increase of phytoplankton, (b) upwelling generally supplies nutrients,
(c) blooms are commonly associate with upwelling or areas with known increases
in nutrients, that extra nutrient supply is required to initiate and maintain
a bloom. However, phytoplankton biomass off Leigh is not normally limited
by nutrients (see series of papers by Taylor et al) and there was no evidence
for any extra supply in 1993.
It is also widely assumed that normal 'spring blooms' not only require
a winter resupply of nutrients, but also the onset of stratification to
'hold' the phytoplankton. This has recently been challenged on the northeast
coast of USA, and stratification would have been impossible at Leigh during
August and September 1992.
5. Coincident and subsequent biological anomalies
Simon Hooker: (2 previous years observations)
(i) Mass mortality (100%) of a single isolated patch (20x1m) of pips
Paphies
(Amphidesma) australe, in Whangateau Harbour, Feb 1993.
(ii) Several occasions when all pipis observed were closed and non-feeding
for several days at a time in the Whangateau Harbour entrance, Jan-Feb
1993.
Others: (1 previous years observation)
(i) mass mortality of juvenile scallops, Pecten novaezelandia,
(appr 35-50mm) in east Great Omaha Bay, around Jan-Feb 1993
(ii) mass mortality of Dosinia ?subrosea in Omaha Bay (both
sides), Jan-Feb 1993
(iii) Starfish disintegration: a low proportion of a scattered population
of Ophidaster luidea? had multiple separated arms, Omaha Bay, Jan-Feb
1993.
W J Ballantine and various newspaper reports - local mass mortalities
of 'shellfish'
(i) Dosinea subrosea - Te Muri, Jan 1992
(ii) grazing molluscs, kina and whelks - Waipu Cove, 5 Feb 1993
Chris Battershill: sponges (comparison with observations in 1980s)
(i) heavy mortality of sponges, especially Callyspongia (near
100%)
(ii) kelp mortality in 1983
Floor Anthoni (diving locally since 1970s), the earliest reports
of:
(i) early kelp sickness and subsequent death, patterned by depth and
geographic locality
(ii) sponge 'shut down' and later death and disappearance, and similar
effects on other fauna.
Russ Babcock (new staff member) and Russell Cole (5+ previous
years observations) (also compared to marine reserve survey 1977-78, see
Ayling, 1978 and Ayling et all, 1981):
Analysis of video transects in the marine reserve (in January
1993) showed earlier and continuing loss of kelp plants (Ecklonia radiata)
from the deeper parts of the bed upwards (demonstrated by the sequential
loss of canopy, stipe, holdfast and bare space). The deeper part of the
kelp beds had 100% mortality and middle parts approximately 50%, throughout
most of the marine reserve. Back calculation (estimating lag time) suggests
a cause pre-December. Second survey in April 1993 showed continuing progressive
mortality into shallower depths, although heavy recruitment has already
occurred in the deeper 'vacant' spaces.
Patchy mortality recorded (in may) from Great Barrier's west coast,
Little Barrier and Whangarei Heads. No sign of mortality on open coast
further north to Cape Karikari, but unconfimred reports of patchy mortality
at Spirits Bay. Similar events associated with cold water reported for
a laminarian kelp in Japan.
P J Smith (MAF), F H Chang (NZOI) and L MacKenzie
(Cawthron):
Have supplied a copy of their draft report for the Royal Society's
Shellfish Toxin Workshop, entitled 'Toxic phytoplankton and algal blooms,
Summer 1992-93'. This collates information for the whole country, mentioning
the Hauraki Gulf events of 1992, but concentrating on events of 1993.
Department of Conservation: reports of high little blue penguin
mortalities in Northland.
Maurice Miles, (North harbour Health): Orewa coughing outbreaks
and sampling for Gymnodinium breve.
6. Probability, pattern and prediction
The question that
unites all aspects of these events, in both theoretical and practical terms
is: When are such events likely to occur? It is also the most important
question from most viewpoints. Biologically we wish to know whether such
events occur sufficiently often to form part of the 'normal' patterns,
are a rarer but still significant disruption of the 'normal' or are very
rare, and hence generally unimportant phenomena. Essentially the same distinctions
are relevant to climatologists, resource managers and economists.
We do not have sufficient direct evidence to answer this question adequately
for any aspect, even those concerning the physical climate anomalies. For
most of the biological aspects there is little or no direct evidence on
frequency distribution in time.
Consider as an example the SST anomalies, for which we have 26 years of
comparable data, and using the term 'return period' borrowed from
engineers concerned with flooding and similar phenomena. If the data are
treated as single set normally distributed (i.e. the form a normal curve)
then September 1992 sea temperatures (which had an anomaly of 3.2 standard
deviations (SD) from the mean) would have a return period greater than
700 years. Even the 6 months April-September 1992 (SD 2.4) would, if treated
as part of a unimodal non-skewed normal distribution, have a return period
of 120 years.
There are, however, at least two reasons for not accepting this analysis.
First, 'normal' distributions were originally derived to describe biological
phenomena within populations and there is no good reason to presuppose
that they apply to phenomena such as climate anomalies. Indeed, despite
much use of statistics based on normal curves (whether transformed or otherwise)
there is growing doubt that they are generally applicable. Professor G
Austin, Geophysics, Auckland (personal communication) refers to 'chaotic'
distributions, which have a spike in the centre and concave side slopes,
giving proportionately more outlying values.
Second, there is reasonable evidence to suggest that the data should not
be treated as a single set. The strong tendency of the SST anomalies to
follow trends in the El Niño/ Southern Oscillation patterns indicates
some form of bimodality. If we adopt the simple case of treating years
separately depending on whether the SOI is positive or negative, the return
period for the April-September 1992 SST anomalies falls from 120 years
to 18 (SD 2.1 from 13 data points.). since SOI is a crude and remote index
for the purpose, it is likely that the return period could be lower still.
These return period calculations are only useful in showing the critical
importance of understanding the patterns of the phenomena. Unless we can
gain some confidence on nature of the patterns we are dealing with, we
can place no reliance on the calculations.
Tf the matter dealt with bench top experiments or easily repeated observations,
teplication (more data ) would be the simple answer to these problems.
But these data points are years. Another 20-40 years of date would be required
to distinguish even the major patterns in statistical terms. Since important
decisions (including social, economic and scientific ones) must and will
be made on much shorter time scales, we are obliged to consider what, if
anything, can be done to assist these.
there are possibilities which include:
'Pattern matching' across different fields of study. It is now being recognised
that scale matching (time and/or space) with physical features is an important,
indeed necessary prerequisite for many biological phenomena in the oceans
(see Mann and Lazier, 1991). The use of satellite data (bothe SST and colour
scanning) would be especially helpful in this respect.
The use of 'proxy data' which can be obtained now, but refers to the past
and can be used to determine, at least, pattern types. Examples include
sediment core analysis.
Connecting to existing longer-run data (for other regions, related parameters,
etc.) by cross-correlations. L Paul did this some years ago connecting
Leigh SST to Leigh air temperatures, and these to Auckland air temperatures
and then the results to snapper recruitment.
It follows from the above that the possibilities of direct causal connections
between the aspects of the 1992-93 events, while interesting, are less
important than whether there are any types of interaction. If there are,
then different aspects can be used as signals for others and pattern matching
becomes highly informative.
The present analysis merely indicated that the return period of the 1992-93
events probably lies in the 10-100 year order of magnitude, and that the
most reasonable guess is that it falls in the biologically critical region
of 15-25 years, i.e. where such events would be important structuring disturbances.
7. Conclusions
The magnitude of these events (geographically, temporally, ecologically
and economically) requires, as a minimum, that we search for a reasonable
level of knowledge on their frequency of occurrence.
The scarcity of data, especially long-term monitoring, will make this difficult.
Even simple routine measurements (e.g. weekly chlorophyll concentrations)
would have improved our understanding considerably.
There is no scientific agreement on how routine marine phytoplankton samples
should be obtained and analysed for general comparability (i.e. for use
as standards, signals and proxy data). Until this confusion is resolved,
useful monitoring of phytoplankton is unlikely to be implemented. The primary
problem is not suitable technology, economic cost or political will but
a scientific consensus that such data is required.
The same points apply to other marine ecological or marine climate data.
References
Anthoni, J F, 1992: The disappearing fish act. Dive Log,
Dec/Jan 1993:13 p31.
Anthoni, J F, 1993: Letter to Editor. Dive Log. Feb/March
1993:14 p7
Ayling, A M 1978: Marine Reserve Survey. Leigh Laboratory
Bulletin 1. Univ Auckland 98pp
Ayling A M, Cumming A and Ballantine W J 1981: Map of shorre
and subtidal habitats of the Cape Rodney to Okakari Point Marine Reserve,
north Island, New Zealand in 3 sheets, scale 1:2000. Dept of Lands
& Survey, Wellington.
Australian Bureau of Meteorolgy: Monthly Climate monitoring Bulletin
(includes world maps of sea surface temperature (SST) and SST anomalies
plus indices such as the Southern Oscillation index, and detailed data
tor the Australian region.
Ballantine W J, 1992: A biologist's view of marine climate.
Weather and Climate (Journal of the NZ Meteorological Scociety) 12:32-39
Booth W E and Sonergaard M, 1989: Picoplankton in the Hauraki
Gulf. NZ J Mar Freshw Res 23:69-78
Durbin E G, 1969: Phytoplankton ecology and productivity
in the Whangateau Harbour. Unpubl MSc thesis, Botany Dept./ Univ Auckl.
175pp
Evans J: annual Leigh Climate Report. Leigh Lab Bull,
Univ auckland.
Harris T F W, 1985: North Cape to East Cape: aspects of the
physical oceanography. 179pp. Physics Dept and Leigh Lab, Univ Auckland.
Haywood A J, 1993: Natural particles in coastal seawater.
Unpubl MSc thesis Env Science, Univ Auckl. 163pp
Heath R A, 1985: A review of the physical oceanography of
the seas around New Zealand - 1982. NZ J Mar Freshw Res 19:79-124
Maddock L and Taylor F J, 1984: The application of multivariate
statistical methods to phytoplankton counts from the Hauraki Gulf, New
Zealand. NZ J Mar Freshw Res 18:309-322
Mann KH and Lazier J R N, 1991: Dynamics of marine ecosystems:
biological-physical interactions in the oceans. Blackwell Sci Publ,
Boston. 466p
New Zealand Herald, daily newspaper. Various reports Dec 1992
onwards. Auckland
NOAA/NWS/NMC monthly Climate Diagnostics Bulletin, Climate
Analysis Centre, Wahsington. Similar to the Australian Climate Monitoring
Bulletin but with more data world-wide.
Rhodes L, Haywood A J,Ballantine W J, MacKenzie L (in prep):
Algal
blooms and climate anomalies off northeast New Zealand, August to December
1992. NZ J Mar Freshw Res.
Smith P J, Chang F H, MacKenzie L, 1993: Toxic phytoplankton
and algal blooms, summer 1992-93. Unpublished paper for the shellfish
toxin workshop. 9pp.
Taylor F J, 1973: Phytoplankton and nutrients in the Hauraki
Gulf approaches. In Oceanography of the South Pacific, 1972. Ed R Fraser,
NZ national Commission for UNESCO, Wellington. p 485-492.
Taylor F J, 1974: A preliminary check-list of microalgae
other than diatoms and dinoflagellates from New Zealand coastal waters.
J Roy Soc NZ 4:395-400
Taylor N J and Taylor F J, 1985: Hydrology and changes in
the nutrient and phytoplankton levels in Goat Island Bay, northern New
Zealand. Int Revue ges Hydrobiol. 86:183-191
Taylor F J, Taylor N J and Walsby J R, 1985: A bloom of the
planktonic diatom Cerataulina pelagica off the cast of northeastern New
Zealand in 1983 and its contribution to an associated mortality of fish
and benthic fauna. Int Revue ges Hydrobiol. 70:773-795
 The bloom commenced
off the Leigh Laboratory at the end of August. It remained intense through
November and declined to more normal levels and composition in December.
The bloom commenced
off the Leigh Laboratory at the end of August. It remained intense through
November and declined to more normal levels and composition in December.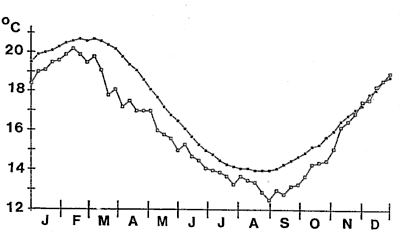 Sea surface temperatures
(SST) were at their lowest when the bloom commenced but solar radiation
had increased markedly some two weeks before.
Sea surface temperatures
(SST) were at their lowest when the bloom commenced but solar radiation
had increased markedly some two weeks before.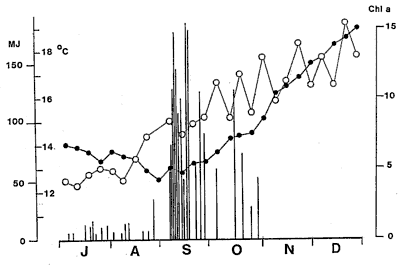 While the precise
triggers for the increase of particular species involved in the bloom and
the limiting factors affecting their subsequent abundance are likely to
vary both specifically and regionally, the widespread nature of the bloom
and its duration suggest some general forcing factor.
While the precise
triggers for the increase of particular species involved in the bloom and
the limiting factors affecting their subsequent abundance are likely to
vary both specifically and regionally, the widespread nature of the bloom
and its duration suggest some general forcing factor.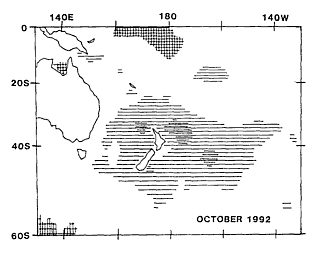
 The question that
unites all aspects of these events, in both theoretical and practical terms
is: When are such events likely to occur? It is also the most important
question from most viewpoints. Biologically we wish to know whether such
events occur sufficiently often to form part of the 'normal' patterns,
are a rarer but still significant disruption of the 'normal' or are very
rare, and hence generally unimportant phenomena. Essentially the same distinctions
are relevant to climatologists, resource managers and economists.
The question that
unites all aspects of these events, in both theoretical and practical terms
is: When are such events likely to occur? It is also the most important
question from most viewpoints. Biologically we wish to know whether such
events occur sufficiently often to form part of the 'normal' patterns,
are a rarer but still significant disruption of the 'normal' or are very
rare, and hence generally unimportant phenomena. Essentially the same distinctions
are relevant to climatologists, resource managers and economists.